O nás
Naše laboratoř se zabývá evoluční protistologií, zajímá nás tedy vznik a evoluce eukaryotické buňky. V současnosti studujeme dvě skupiny protist, Preaxostyla a Euglenida. Zkoumáme jejich diverzitu, molekulární a buněčnou biologii. Zaměřujeme se zejména na semiautonomní organely těchto organismů.

Preaxostyla a Euglenida jsou skupiny bičíkovců patřící pod superskupinu Excavata. Nejvíce nás zajímá evoluce jejich semiautonomních organel: vysoce redukovaných či zcela ztracených mitochondriálních organel anaerobní skupiny Preaxostyla a sekundárních plastidů fotosyntetických euglen. Výzkum obou těchto oblastí by nám mohl pomoci pochopit původ organel a evoluci jejich struktur, molekulární biologii, transport, targeting, biogenezi, kompozici genomu a biochemické dráhy. Studujeme též fenomén laterárního genového transferu, který hraje důležitou roli v některých těchto procesech.
Preaxostyla
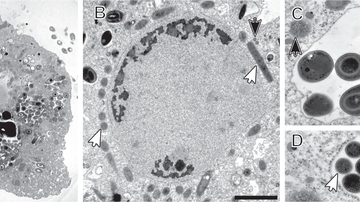
Organismy patřící do skupiny Preaxostyla žijí výhradně v anoxickém prostředí. Jedná se o jednu z nejméně prozkoumaných linií eukaryot. Věříme, že výzkum těchto protist nám může pomoci porozumět procesu reduktivní evoluce mitochondriálních organel. Paratrimastix pyriformis patří v rámci této skupiny mezi bazální, volně žijící organismy. Studiem fyziologických funkcí redukované mitochondrie tohoto organismu se snažíme se objasnit průběh redukce mitochondrií u ostatních preaxostyl. Dále nás zajímá skupina Oxymonadida, kam patří obyvatelé trávicího traktu nejrůznějších živočichů. Náš výzkum druhu Monocercomonoides exilis ukázal, že se pravděpodobně jedná o skupinu, která mitochondrii zcela ztratila.
zaměřujeme se na:
- transkriptomiku a genomiku oxymonád a paratrimastixe
- buněčnou lokalizaci předpokládáných mitochondriálních proteinů
- metabolismus spojený s anaerobiózou a ztrátou mitochondrie
- dráhy pro syntézu železo-sirných center
- symbiózy oxymonád a prokaryot
- diverzitu oxymonád

Euglenida
Euglenida je skupina převážně sladkovodních bičíkovců. Jejich mitochondrie jsou neobvyklé z hlediska struktury i molekulární genetiky. Mnoho euglen má pevnou ale pružnou pelikulu, díky které jsou schopné metabolie, tedy typického euglenoidního pohybu. Eugleny jsou také známé pro heterogenitu ve způsobu získávání energie. Ancestrální a nejrozšířenější je v této skupině heterotrofie (bakteriovorie, eukaryovorie a primární osmotrofie). Jedna z monofyletických skupin, Euglenophyta, získala v průběhu evoluce sekundární zelený plastid, a jako hlavní zdroj energie využívá fotosyntézu. Tento plastid je derivovaný z prasinofytních řas a má tři membrány. I přesto jsou tyto eugleny schopné přežít bez přítomnosti světla díky schopnosti přechodně přepnout na heterotrofii. Tato schopnost zapříčinila vzniku několika osmotrofických druhů s nefotosyntetickým sekundárním plastidem. Rapaza viridis, nedávno objevený mixotrofický druh, vyžaduje k přežití jak fotosyntézu, tak eukaryotickou kořist.
zaměřujeme se na:
- transkriptomiku a genomiku plastidů euglen a příbuzných řas
- laterální genový přenos spojený se vznikem plastidu
- plastidovou proteomiku a proteinový import u Euglena gracilis
- analýzu lipidů chloroplastu euglen
- hledání organismů příbuzných předkovi plastidu euglen pomocí enviromentální sekvenace
- diverzitu heterotrofních euglenidů

V současné době probíhá výzkum v těchto směrech:
Buňky bez mitochondrií
- V roce 2016 se nám podařilo odhalit, že prvok Monocercomonoides exilis zcela postrádá mitochondrii. Jedná se o vůbec první eukaryotickou buňku, které tato organela prokazatelně chybí. Nyní se snažíme proniknout hlouběji do tajů funkcí takových buněk, zejména nás zajímá funkce syntézy železo-sirných center, což je proces, který s mitochondrií velice úzce souvisí.
- Rádi bychom také charakterizovali některé přenašeče elektronů účastnící se redoxních reakcí, jak u těchto amitochondriátů, tak jejich blízkých příbuzných.
- Zajímá nás také diversita oxymonád a rádi, bychom získali, co možná nejkompletnější fylogenetický strom oxymonád, a proto stále lovíme a sekvenujeme oxymonády z prostředí, kde se vyskytují.
- Chceme také použít amitochondriální buňky k „evolučním“ pokusům simulujícím situaci, kdy v buňkách vznikala mitochondrie. Zajímá nás například, co se stane, když smícháme cytosolický extrakt amitochondriálních oxymonád s izolovanými mitochondriemi jiných organizmů. Které proteiny budou importovány do izolovaných mitochondrií?
- Oxymonády nejsou možná jediné organizmy bez mitochondrií. Ve spolupráci s kolegou Ivanem Čepičkou máme na mušce další dvě skupiny prvoků, které bychom rádi prověřili na přítomnost této organely. Jednou z nich jsou améby z rodu Pelomyxa a druhou bičíkovci rodu Retortamonas.
Plastidy krásnooček
- Je naprosto jasné, že plastid zelených krásnooček vznikl sekundární endosymbiózou se zelenou řasou. Dokonce se ví, že tato řasa pocházela z evolučního okolí rodu Pyramimonas. Vzhledem k tomu, že první větve fylogenetického stromu zelených krásnooček jsou tvořeny mořskými zástupci, se domníváme, že k této události došlo v moři. Stáří této události neznáme, ale je zřejmé, že se jedná o mladší endosymbiózu, než je většina ostatních.
- V našem výzkumu se snažíme prozkoumat podrobnosti této události. Zajímá nás, jak se vyvíjely genomy plastidů, a proto sekvenujeme plastidové genomy zelených řas a krásnooček, které se větví co nejblíže endosymbotické události. Ukazuje se, že diverzita mořských zelených i nezelených krásnooček je zmapována jen povrchně, a tak ji chceme zkoumat důkladněji a doufáme v objev nových linií.
- Dále nás zajímá, jak plastidy krásnooček fungují. Zejména se snažíme zjistit, jaký je původ plastidových membrán. Mají totiž jen tři membrány oproti předpokládaným původním čtyřem takže, která se ztratila? Nejasný je také způsob transportu proteinů z cytosolu přes plastidové membrány. Máme vytipovaných několik málo proteinů, které by se toho mohly účastnit, ale nemáme potvrzenu jejich lokalizaci a už vůbec netušíme s jakými dalšími proteiny spolupracují.
- Rádi bychom se také naučili krásnoočka transfekovat cizorodou DNA a potlačovat expresi jejich genů metodami RNAi.
Cytoskelet protist
- O proteinovém složení neaktinového a netubulinového cytoskeletu prvoků, tedy různých druhů intermediánních a žíhaných fibril, se toho ví velmi málo. Přitom by tato znalost mohla umožnit homologizovat některé morfologické struktury napříč vzdálenými skupinami a odpovědět například na otázku, jak vypadal předek eukaryot (LECA). Domníváme se, že metodika uzrála do stadia, kdy je možné si začít takové otázky klást. Předběžné pokusy ukazují, že jsme schopni připravit obohacenou frakci cytoskeletu oxymonád. Následná proteomická analýza by mohla prozradit mnohé o jeho složení.